4. Einführung in die Ultrastruktur und den Stoffwechsel der Bakterien
4.1. Vergleich der strukturellen Organisation der Organismengruppen
4.1.1.
Unterschiede
zwischen Pro- und Eucaryonten.
Bakterien
vertreten die Procaryonten. Per Definition handelt es
sich dabei um Lebensformen, deren zelluläre Organisation ohne einen Zellkern
auskommt, d.h., das Erbgut liegt im Cytoplasma vor.
Höhere Lebewesen sind gekennzeichnet durch das Vorhandensein eines Zellkerns in
ihren Zellen (wobei es hiervon Ausnahmen gibt, so sind z.B. die Erythrocyten kernlos). In diesen Kernen liegt die gesamte
Erbinformation in Form von Chromosomen vor. Neben dieser namengebenden
Eigenschaft, die Pro- und Eucaryonten unterscheidet,
gibt es noch einige grundsätzlich wichtige Merkmale, die in der unten stehenden
Tabelle zusammen gefasst sind.
Tabelle 6: Wesentliche
Unterschiede zwischen Pro- und Eucaryonten.

Die
strukturellen Unterschiede zwischen Eucaryonten und Procaryonten schlagen sich in der Möglichkeit nieder,
parasitische Procaryonten mit selektiv toxischen
Antibiotika zu bekämpfen, während sie den Makroorganismus, ihren Wirt,
besiedeln. Die selektive Toxizität der
Antibiotika wird dadurch gewährleistet, dass in solchen Fällen, in denen es
Homologien zwischen (molekularen) Strukturen der
Bakterien und den Zellen des Wirtes gibt, die entsprechenden homologen
Entwicklungen einen genügend großen Unterschied aufweisen (verg. z.B. die Ribosomen : Ribosomen gibt es zwar bei Pro- und Eucaryonten, ihre Funktion ist im
Prinzip identisch, in der Feinstruktur unterscheiden sie sich jedoch zwischen
beiden Zelltypen sehr deutlich).
Auf solche Unterschiede, welche die
selektive Toxizität der Antibiotika gewährleisten, wird im Zuge der Vorlesung
immer wieder hingewiesen. (vergl. auch die Hinweise zur Resistenz gegen Antibiotika im Kapitel 5 Genetik).
4.1.2.
Unterschiede
zwischen Procaryonten und Viren
Viren
stellen eine Sonderform in der Natur dar. Obwohl sie in der Lage sind, z.T. lebensbedrohliche Erkrankungen auszulösen, handelt es
sich doch streng genommen um unbelebte Partikel, das sie zu einer
eigenständigen Vermehrung wie auch zu einem selbstständigen Stoffwechsel nicht
in der Lage sind. Auf die Viren wird in Kapitel 6 näher
eingegangen, so dass hier nur die wesentlichen Unterschiede zu den Porcaryonten aufgelistet werden.
— Typ des Erbguts (Viren weisen i.d.R.
nur einen Typ Nukleinsäure (RNA oder DNA) auf, nur in Ausnahmen beide Typen)
— kein eigener Stoffwechsel
— keine zelluläre Organisation
— nicht selber vermehrungsfähig
à Viren sind keine
Lebewesen sondern infektiöse Partikel.
4.2.
Struktur und
Funktion der bakteriellen Zellwand
Die meisten Bakterien sind gegenüber dem umgebenden
Milieu durch eine Zellwand abgeschottet. Funktionen dieser Zellwand sind die
Formgebung (Stäbchen, Kokken etc.) und der Schutz gegenüber schädigenden
Substanzen. Die bakterielle
Zellwand ist in ihrer molekularen Struktur (Grundbaustein: Murein,
Peptidoglycan) in der belebten Welt einzigartig. In
dieser Zusammensetzung gibt es die Zellwand nur bei den Bakterien. Damit wird
die bakterielle Zellwand zum bevorzugten Ziel für antibiotische
Substanzen! Das Penicillin, dessen Entdeckung den Beginn des antibiotischen Zeitalters markierte, wirkt auf die Zellwandsynthese
der Bakterien ein (Abb. 10), ebenso wie eine mittlerweile große Anzahl
Strukturverwandter Antibiotika aus der Gruppe der Penicilline und Cephalosporine. Die Zellwand der Pflanzenzellen
(Grundbaustein: Cellulose & Pectin) und
Pilzzellen (Grundbaustein: Chitin) unterscheidet sich von der Bakterienzellwand
grundsätzlich.
Im
Gesamtsystem der Bakterien weist die Zellwand Variationen desselben Themas auf.
So gibt es neben zellwandlosen Bakterien (z.B. Mykoplasma ssp.)
zwei grundsätzliche Typen der Zellwand, welche man auch mit Färbemethoden die
in der Routinediagnostik verwendet werden, unterscheiden kann. Es handelt sich
hierbei um die Zellwand der Gram positiven und die der Gram negativen
Bakterien. Die sogenannte Gram-Färbung erlaubt die
Unterscheidung beider Zellwandtypen im Mikroskop und erlaubt damit im
Routinelabor auch eine erste Einteilung isolierter Bakterien.
4.2.1.
Aufbau der Gram positiven Zellwand
(Beispiele: Staphylococcus aureus, Streptokokken
, Milzbranderreger)
Grundbaustein ist das Murein,
ein Polymer aus N-Acetylmuraminsäure (NAc) und N-Acetylglucosamin (NAcGlc). An den NAc-Resten sind i.d.R. fünf Aminosäuren gebunden (daher die chemische
Bezeichnung Peptidoglycan):
(L-Ala▬D-Glu▬L-Lys▬D-Ala▬D-Ala)
oder
(L-Ala▬D-Glu▬D-amino-Pimelinsäure▬D-Ala▬D-Ala)
Über diese Aminosäurereste erfolgt mit Hilfe von Pentapeptiden (5 Glycinreste)
eine Quervernetzung der Mureinstränge unter Abspaltung des Endständigen D-Ala (Abb. 10). Diese quervernetzte Struktur ist von
außerordentlicher Festigkeit und verleiht der Bakterienzelle ihre äußere Form.
In der Gram positiven Zellwand findet man einen
„mächtigen“ Polylayer aus Mureinsträngen als
Hauptbestandteil der Zellwand (siehe auch Abb. 12). Gram positive Bakterien
erscheinen im Lichtmikroskop blauviolett. Es gibt jedoch einige Bakterien wie
z.B. die Mitglieder des Genus Mycobacterium,
die zwar eine im Aufbau Gram positive Zellwand aufweisen, sich jedoch auf Grund
feiner struktureller Unterschiede nicht mit der Gramfärbung sondern nur mit
speziellen Färbemethoden anfärben lassen.

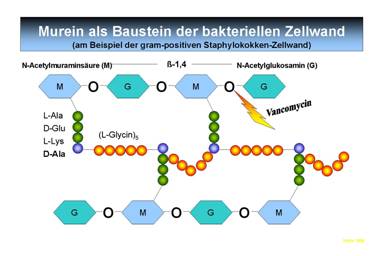
Abb. 10: Grundbausteine und vereinfachte Schritte der Synthese der
bakteriellen Zellwand bei Gram positiven Erregern. Die untere Abbildung zeigt
die Quervernetzung des Peptidoglycans unter
Abspaltung des endständigen D-Ala Restes zur fertigen
Zellwand. Angriffspunkte für Antibiotika sind markiert. Vancomycin
hemmt die glykosidische Bindung der Einzelzucker zum
Polymer während Penicillin den Transpeptidierungsschritt
während der Quervernetzung hemmt. Es wird deutlich, dass Antibiotika ihre
Wirkung nur auf Teilungsaktive Bakterien ausüben können. Eine bereits
bestehende Quervernetzung wird durch Antibiotika nicht mehr gestört. Der
Angriff bspw. des Penicillins erfolgt auf spezifische Enzyme, welche die Transpeptidierung katalysieren.
4.2.2.
Aufbau der Gram negativen Zellwand
(Beispiele: Neisseria
meningitidis , E. coli, Salmonellen u.a.)
Grundbaustein ist auch hier Murein,
allerdings erfolgt die Quervernetzung i.d.R. ohne
Pentapetide direkt über die seitenständigen Aminosäuren des NAc
(Abb. 11). Die Zellwand besteht nur aus einem Mono- oder Bilayer.
Darüber befindet sich als Abschluss nach außen eine äußere Membran
(Charakteristikum der Gram negativen Zellwand). Nach Gramfärbung erscheinen
Gram negative Bakterien im Lichtmikroskop rötlich. Auch in diesem Fall gibt es
jedoch Abweichungen im Feinbau der Gram negativen Zellwand. So lassen sich z.B.
die strukturell Gram negativen Spirochäten (u.a. die
Erreger der Syphilis, Treponema pallidum) i.d.R. nicht nach Gram anfärben.

Abb. 11: Aufbau der bakteriellen Zellwand bei Gram negativen
Erregern. Beachte die im Vergleich zu Gram positiven Bakterien (Abb. 10)
unterschiedliche Art der Quervernetzung des Peptidoglycans.
Angriffspunkte für Antibiotika sind markiert (anders als Vancomycin
oder Penicillin ist das Lysozym eine sehr weit
verbreitete antibiotische Substanz, welche man z.B.
in Tränenflüssigkeit nachweisen kann).
Die
Synthese des Mureins kann in verschiedenen Stadien durch Antibiotika gestört
werden (vergl. Abb. 10/Abb. 11). Die bereits
bestehenden Zellwandstrukturen werden von Antibiotika in der Regel nicht mehr
zerstört. Die Gruppe der Penicilline und Cephalosporine
gehört z.B. zu den Substanzklassen, die in die Zellwandsynthese der Bakterien
eingreifen.
Gram
positive und –negative Erreger unterscheiden sich nicht nur in der Art der
Quervernetzung des Mureins sondern auch in einigen grundsätzlichen Punkten im
groben Aufbau der Zellwand (Mächtigkeit der Mureinschicht; äußere Membran bei
Gram negativen Bakterien; siehe Abb. 12). In die Zellwand eingelagert oder
aufgelagert sind unterschiedliche Proteine, Polysacharide etc., welche zum Teil
wichtige Rollen in der Pathogenese von Erkrankungen spielen.


{kind=link}
{kind=link}
Abb. 12: Beispiele für den Aufbau der Gram
negativen Zellwand (links am Beispiel der Meningokokken) bzw. der Gram
positiven Zellwand (rechts am Beispiel der Streptokokken). Die in der Membran
liegenden Proteine bzw. die der Membran oder der Zellwand aufgelagerten
Polysaccharide und Lipopolysaccharide
(Gram negative) bzw. Lipoteichonsäure (Gram positive)
stellen Pathogenitätsfaktoren der Bakterien dar.